ISOLATION AND PHYSICOCHEMICAL CHARACTERIZATION OF BRUCELLA PHAGES
A. Y. Shaheen1, A. A. Sheikh*1, M. Rabbani1, W. Shehzad2, Z. Abbas3 and M. Maqbool1
1Institute of Microbiology, 2Institute of Biochemistry and Biotechnology, University of Veterinary and Animal Sciences, Sheikh Abdul Qadir Jelani Road Lahore Pakistan; 3Department of Microbiology & Molecular Genetics, Canal Bank Rd, Punjab University New Campus, Lahore, Punjab
* Corresponding Author’s E-mail: ali.ahmad@uvas.edu.pk
ABSTRACT
Bovine brucellosis, caused by Brucella abortus, is an economically significant bacterial disease causing enormous economic losses in developing countries. Due to emerging antibacterial resistance in current use of antibiotics and insufficient immunity by WHO recommended vaccine strategies, it is recommended to cull the positive animals to control the disease. In such circumstances, use of host specific bacteriophages could be an alternate option to control the disease. In present study, brucellaphages were isolated from slurry samples (n=50) of livestock farms. Seven samples were found positive in spot method, while two samples gave the positive plaques of pinpoint size (0.5 mm) with round and clear appearance in plaque assay. Isolated brucellaphages (BaP1 and BaP2) did not produce plaques against Staphylococcus aureus, Streptococcus, Bacillus, Escherichia coli, Salmonella, and Pasteurella multocida. Physicochemical characterization revealed that lytic activity of phages was present up to 60oC which started to decrease at 70oC and maximum stability was between 7 to 9 pH. Exposure of sunlight, normal fluorescent and UV light inactivated these phages within 3 hours, 24 hours and 15 minutes, respectively. Phages become inactivated in 15 minutes when treated with Sodium Dodecyl Sulphate, chloroform, Lysozyme, Proteinase K and EDTA, however, no effect of normal saline, Trypsin and RNAse was observed on brucellaphages. In conclusion, the results have laid the foundation to standardize practical applications of brucellaphages after detailed in-vitro and in-vivo experimental evaluations.
Keywords: Brucellosis, Brucella abortus, brucellaphage, plaque assay, physicochemical characterization
https://doi.org/10.36899/JAPS.2021.5.0328
Published online January 21, 2021
INTRODUCTION
According to economic survey of Pakistan, the livestock sector has ability to grow rapidly but due to prevailing issues of management, farm biosecurity and infectious diseases, it is not growing at high pace. Brucellosis is one of the significant infectious bacterial diseases found in developing countries. Bovine brucellosis mainly caused by Brucella abortus (B. abortus), is a zoonotic disease with an economic importance, worldwide (OIE, 2019).
Host range of B. abortus includes cattle, camels, water buffalo, bison, and elk, while infrequently other species of animals also get infection. In most of the animal species, the disease predominantly affects the reproductive system with concomitant loss in productivity (Manish et al., 2013). Ruminants become chronic asymptomatic carrier of bacteria after their first abortion and they can cause environmental contamination through shedding of bacteria. Consequently, during successive pregnancies, animals frequently shed Brucella in milk and also in uterine secretions (Shang et al., 2002). The challenges in controlling brucellosis include development of compatible relationship of Brucella species among various animal hosts, incubation period; suitable conditions for bacterial survival outside and within the host; long and asymptomatic carrier stages as well as non-availability of proper treatment. As animal husbandry practices are not up to the mark and animal population is increasing rapidly, the control of brucellosis is further getting difficult (Rahman et al., 2006). In Pakistan, with restricted resources, limited veterinary facilities, and due to high cost of dairy animals, eradication of the disease by eliminating all Brucella infected animals across the country is not feasible (Abubakar et al., 2012).
Bacteriophages are bacterial viruses which exclusively infect and replicate inside specific bacterial species and results in killing of targeted bacteria (Wommack and Colwell, 2000; Prestel et al., 2008; Srinivasiah et al., 2008). As, phages attack specific bacteria so, they do not cause resistance in untargeted bacterial species (Principi et al., 2019; Moghadam et al., 2020). The phages kill the bacterial cells by establishing lytic cycle for own replication using metabolic machinery and genes of host bacteria. Recently, phage therapy has arisen as promising alternate to antibiotics as enormous use of antibiotics has resulted in the emergence of antibiotic resistance all over the world (Hyman and Abedon, 2010; Gopalaiah, 2013). Previous studies revealed that brucellaphages were used for bacteriophage-based diagnostics, phage typing and epidemiological studies (Gupta and Saxena, 2017a; Sergueev et al., 2017). Most of the studies had investigated the genetic diversity among previously characterized reference brucellaphages (Tb, Bk, R/C, Wb, Fi, Iz, Pr) but remain limited about successful use of brucellaphages for specific decontamination and antibacterial therapy (Zhu et al., 2009; Flores et al., 2012; Filippov et al., 2013; Farlow, 2014; Tevdoradze et al., 2015; Hammerl et al., 2014, 2017). Only a few investigations have been concerned with the use of specific bacteriophage for prophylaxis and therapy against brucellosis (Chachra et al., 2012; Pandey et al., 2013; Prajapati et al., 2014; Jain et al., 2015; Gupta and Saxena, 2017b; Saxena and Raj, 2018). Conferring literature reviewed, this is the first study of isolation of brucellaphages from Punjab, Pakistan which will concrete a way for its use in several practical applications. Thus, present study was designed for the isolation and characterization of indigenous isolates of bacteriophages against infective B. abortus, which may be available for future application in livestock and dairy farms to control brucellosis.
MATERIALS AND METHODS
Study area: This study was conducted at University Diagnostic Laboratory (UDL), University of Veterinary and Animal Sciences (UVAS), Lahore in the year 2018.
Bacterial strain:Brucella abortus S99, procured from Veterinary Research Institute (VRI), Lahore was used as host strain under strict biosafety precautions as per recommended in OIE (2018) for the isolation of bacteriophages.
Collection and processing of samples: A total 50 sewage drainage water/slurry (semi-liquid mixture of manure) samples were collected, out of which 42 samples were of slurry, collected from 10 livestock farms located in Lahore (n=14), Pattoki (n=8), Hafizabad (n=3), Sargodha (n=2), Okara (n=13) and Bahawalpur regions (n=2), while 8 sewage drainage samples were collected from Lahore (n=6), Pattoki (n=1) and Okara (n=1) regions. The selection of the livestock farms was based on their previous history of brucellosis in dairy animals as tested by Rose Bengal Test (RBT) and Brucella specific ELISA at University Diagnostic Lab, University of Veterinary and Animal Sciences, Lahore. The collected samples were processed using the modification in the method of Chachra et al. (2012). Slurry samples, mixed with normal saline (4:1), were kept on shaking (120 rpm) (Robus Technologies, SI900R) overnight at 25oC to dissociate suspected bacteriophages from organic material present, followed by centrifugation (Eppendorf 5810, Germany) at 4,000 × g for 15 min. In modification, 50 mL double strength (2X) Tryptose Soy Broth (TSB) (Merck Millipore, Germany), 40 mL slurry supernatant and 10 mL broth culture of B. abortus S99, in exponential growth phase (24 hr) having sufficient bacterial turbidity (O. D600=0.8), were added and incubated at 37oC for one week with shaking at 120 rpm. Mixture was centrifuged and filtered sterilized through 0.45 µm filters (Millex® Millipore) for further enrichment (Chachra et al., 2012).
Enrichment of samples: Successive enrichment of three to four times was given to phage filtrates, obtained after preliminary screening of the samples by spot assay, with B. abortus S99 to increase the phage titer. In brief, equal volume of filtrate (25 mL) and (2X) TSB (25 mL) with 1 mL of 24 hr old bacterial culture (exponential phase) were added and incubated at 37oC, for 48-72 hr with shaking at 150 rpm. Then, 10 mL sample was drawn out from the mixture and centrifuged at 8,000 × g, for 15 min. Supernatant was collected and filter sterilized using 0.45 µm and 0.2 µm syringe filters (Millex® Millipore). Filtrate was stored at 4oC for further processing (Texas, 2011).
Processing of enriched cultures: Isolation of bacteriophages was carried out by spot and plaque assays using the double agar overlay technique (Yang et al., 2010). Using this technique, filtrate from enriched cultures was mixed with host bacteria (log phase) in soft agar and was overlaid on hard Tryptose Soy Agar (TSA) plates. Semisolid TSB with 0.65% agarose, (maintained at 45oC in water bath) was used as soft agar for overlaying.
Spot assay: Spot assay was used as screening test for the isolation of bacteriophages. B. abortus S99 culture 200 µL (log phase=24 hr) was mixed in 3-5 mL of soft agar and was spread evenly over TSA media plate. Filtrate (15 µL) was spotted onto plates having lawn of host culture and incubated at 37oC for 48 hr (Yang et al., 2010).
Plaque forming assay: Plaque forming assay was used for purification of bacteriophages. 100 µL filtrate and 200 µL of host culture were mixed in 3-5 mL soft agar and were overlaid onto TSA plates followed by incubation at 37oC for 48 hr. Before, adding into soft agar, filtrate and bacterial host culture were given varied adsorption time (15, 30, 60, 90 and 120 min) by incubating the mixture at 37oC. To enhance the recovery of plaques, 30 µL CaCl2 and MgCl2 (each at a concentration of 0.05 M) were also added in soft agar (Yang et al., 2010). Results for all the experiments included morphology of the plaques, host range and physicochemical characterization were compared with the positive control brucellaphage strain, Tbilisi (Tb) phage, obtained from Felix d’ Herelle Reference Center for Bacterial Viruses (Laval University, QC, Canada).
Elution of brucellaphages: The plate having well isolated plaques was selected for the elution of bacteriophages. Elution was carried out by pouring 2 mL of Saline Magnesium (SM) buffer (100 mM NaCl, 25 mM Tris-HCl pH 7.5, 8 mM MgSO4, 0.01% (w/v) gelatin) over the top layer (semisolid) and gently agitated the layer with the sterilized wire loop to release the phages from the semisolid agar. The SM buffer was aspirated and centrifuged at 5,000 × g to remove the agar pieces. Subsequently, supernatant was filtered through 0.22 µm filters to remove the bacteria and elute the phage in SM buffer (Chachra et al., 2012).
Purification of plaques: Bacteriophages were purified by picking the well isolated single plaque with sterilized micropipette tip, propagated in 10 mL broth of Brucella culture overnight and growing through double agar overlay technique. Sterile filtered phage lysates were diluted in TSB to six different dilutions (10-1-10-6). The process was repeated for 2-3 times until homologous plaques were obtained (Hamza et al., 2016).
In vitro assessment of broad-spectrum lytic activity of brucellaphages: In vitro lytic spectrum of the purified bacteriophages (n=2 positive isolates) (BaP1 and BaP2) was checked against various heterologous species of bacteria of veterinary significance viz. Staphylococcus aureus (ATCC® 23235TM), Streptococcus sp. (ATCC® 9884TM), Pasteurella multocida (ATCC® 43137TM), Bacillus sp. (ATCC® 27860TM), E. coli (ATCC® 25922TM), and Salmonella spp. (ATCC® 35664TM) (Chachra et al., 2012). In vitro lytic activity of brucellaphages against homogeneous species was not tested due to non-availability of B. abortus strains.
Physicochemical characterization of brucellaphages: Purified bacteriophages (BaP1 and BaP2) were characterized for their physicochemical properties by studying the following parameters:
Effect of different temperatures on brucellaphages: Thermal stability of (BaP1 and BaP2) was checked by incubating phage suspensions in TSB (pH=7.0) at various temperatures (25, 37, 45, 60, 70, 80, 90°C) for one hour. The incubated phages were individually mixed with host bacteria (log phase) and phage titers were evaluated by double agar overlay method. Phage-free suspensions and bacterial-free suspensions were used as controls incubated under the same conditions as the phage-bacterial suspensions (Hamza et al., 2016).
Effect of pH on brucellaphages: The effect of different pH values on (BaP1 and BaP2) stability was determined by incubating phage suspensions in TSB adjusted in steps of 1 pH unit from pH 2 to 9 for one hour at 37°C. Subsequently, phage titers were checked by double agar overlay method described above (Hamza et al., 2016).
Effect of storage temperatures on brucellaphages: To determine the most suitable storage temperature, stability of (BaP1 and BaP2) was checked by storing at 4, -20 and -80°C for 1, 6 and 12-months period. TSB (4oC) and glycerol (-20 and -80oC) stocks were prepared for the storage of brucellaphages (Hamza et al., 2016).
Effect of light on brucellaphages: 100 µL of (BaP1 and BaP2) (106 pfu/mL) was exposed to UV light, sunlight and normal fluorescent tube light for a period of 15, 60, 120 min, 3 hr and 24 hr. Change in pfu was observed by adding 200 µL of freshly grown B. abortus culture and treated phages in cooled molten semisolid TSB and overlaid on TSA plates. Plates were incubated at 37°C for 48 to 72 hr aerobically (Chachra et al., 2012).
Effect of enzymes and chemicals on brucellaphages: 100 µL of (BaP1 and BaP2) (106 pfu/mL) was treated with 20 µL of enzymes viz. Proteinase K (20 mg/mL), Trypsin (250 µg/mL), Lysozyme (20 mg/mL), and (RNAse 10 mg/mL). Similarly, phages were treated with equal volume of Sodium Dodecyl Sulphate (SDS) (10%), Normal Saline Solution (NSS), chloroform and EDTA (0.01M) for 15, 30 and 60 min. Change in pfu was observed by adding 200 µL of freshly grown B. abortus culture and treated phages in cooled molten semisolid TSB and overlaid on TSA plates. These plates were incubated at 37°C for 48 to 72 hr aerobically (Chachra et al., 2012). The data were analyzed statistically using McNemar Test and, one-way Analysis of Variance (ANOVA) with 95% confidence interval using Statistical Package for Social Sciences (SPSS) version 23.0.
RESULTS
Isolation of brucellaphages for B. abortus: Out of 50 sewage water/slurry samples processed, 7 samples (14%) were found positive in spot assay (screening), while 2 out of 7 screened samples produced plaques in plaque assay for brucellaphages (BaP1 and BaP2). Both phages have identical plaque morphology (round, clear and pinpoint plaques having (0.5 mm) diameter after 48 hr of incubation at 37oC. However, they differ in isolation source/sample nature i.e. BaP1 was isolated from slurry of livestock farm and BaP2 from sewage sample. Pinpoint plaques of BaP1 were observed on plates after 48 hr of incubation (Fig. 1). Optimum adsorption time for the brucellaphages was determined to be 120 min in this study as maximum plaques (4.5 × 10-6 pfu/mL) were recovered in 120 min, when various adsorption time (15, 30, 60, 90 and 120 min) were given before adding into soft agar. Plaque morphology of brucellaphages BaP1 and BaP2 was similar to that for Tb phage. Statistically non-significant difference 0.062 (p>.05) was observed in spot and plaque assay results through McNemar Test.
Figure 1. Presence of pinpoint, round and clear plaques indicated the presence of brucellaphage (BaP1), against Brucella abortus S99
Host range of brucellaphages: Broad spectrum lytic activity of brucellaphages (BaP1 and BaP2) by plaque assay against other heterogeneous bacterial species indicated that they were unable to lyse Bacillus, Staphylococcus aureus, Streptococcus, Pasteurella multocida, E. coli and Salmonella. However, positive lytic spots were observed against Staphylococcus aureus and Bacillus in spot assay.
Physicochemical properties of brucellaphages:
Thermal stability of brucellaphages: BaP1 and BaP2 phages found to withstand temperature up to 60oC, but their viability starts decreasing when exposed to 70oC. While, they were inactivated completely in 1 hr exposure beyond 70oC (Fig. 2). Tb phage had stability up to the 80oC but beyond 80oC it was inactivated in 1 hr exposure. Statistically non-significant difference (p>.05) was observed in phage viability at 25, 37, 45 and 60oC, however, phage viability was significant (p=0.00) at higher temperatures.
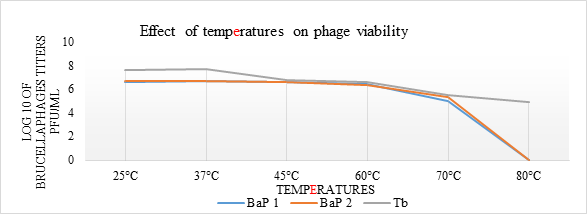
Figure 2. Graphical representation of thermal stability of brucellaphages. Phage suspensions in TSB (pH=7.0) were incubated for one hour at adjusted temperatures. Post treatment viability was assessed through double agar overlay technique. Log10 values of phage titers (y-axis) were plotted against various temperatures (x-axis) for phage isolate 1 (BaP1), phage isolate 2 (BaP2) and positive control Tbilisi phage (Tb).
Effect of pH on brucellaphages: Both phages showed maximum stability between pH 7 to 9 but between acidic pH (2 to 4) their viability was decreased in 1 hr exposure (Fig. 3). Statistically significant difference (p=0.00) was observed in phage viability at acidic and alkaline pH values.
Effect of storage temperatures on brucellaphages: Both BaP1 and BaP2 phages stocks stored in TSB showed maximum stability at 4oC as no loss in phage titer was observed up to 12 months. But loss of phage titer to 50% and 100%, in glycerol stocks, was observed at -20 and -80oC, respectively after 12-months period (Fig. 4). Effect of all three storage temperatures (4oC, -20oC and -80oC) showed significant (p<.05) response on the survival of brucellaphages.
Effect of light on brucellaphages: BothBaP1 and BaP2 phages showed considerable stability, with slight reduction of (5%) and (19%) in titers after 1 hr exposure in fluorescent and sunlight, respectively. But exposure of 2 hr in fluorescent and sunlight reduced viability of phages up to 7% and 32%, respectively. However, UV light killed these brucellaphages within 15 min of exposure (Fig. 5). In comparison of effect of lights, significant difference (p<.05) was observed between effect of UV light with fluorescent and sunlight.
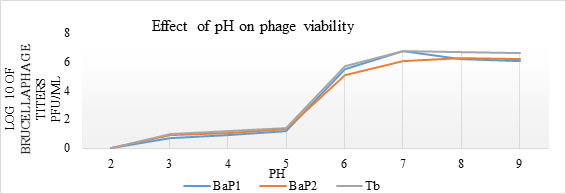
Figure 3. Graphical representation of stability of brucellaphages at acidic and basic pH values. Phage suspensions in TSB adjusted in pH range 2 to 9 were incubated for one hour at 37°C. Post treatment viability was assessed through double agar overlay technique. Log10 values of phage titers (y-axis) were plotted against different pH values (x-axis) for phage isolate 1 (BaP1), phage isolate 2 (BaP2) and positive control Tbilisi phage (Tb).
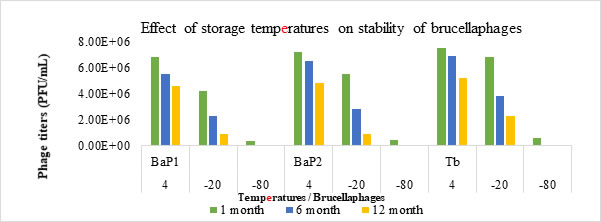
Figure 4. Pictorial representation of effect of storage temperatures on stability of brucellaphages. Phage suspensions were incubated at different storage temperatures for 1, 6- and 12-month period. Post treatment viability was assessed through double agar overlay technique. Phage titers (PFU/mL) (y-axis) were plotted against effect of storage temperatures (4oC, -20oC, -80oC) (x-axis) for phage isolate 1 (BaP1), phage isolate 2 (BaP2) and positive control Tbilisi phage (Tb).

Figure 5. Pictorial representation of effect of different lights on stability of brucellaphages. Phage suspensions were incubated in different lights for time durations of 15, 60- and 120-min. Post treatment viability was assessed through double agar overlay technique. Phage titers (PFU/mL) (y-axis) were plotted against effect of Ultraviolet light (UV), Fluorescent light (F) and Sunlight (S) (x-axis) for phage isolate 1 (BaP1), phage isolate 2 (BaP2) and positive control Tbilisi phage (Tb).
Effect of enzymes and chemicals on brucellaphages: Both BaP1 and BaP2 phages showed survival rates of 19.03% and 21.6% when treated with Lysozyme and Proteinase K, respectively for 15 min. But survival rate of the phages was 100% when exposed to Trypsin and RNase (Fig. 6). No effect on viability of brucellaphages was observed after treatment with NSS and EDTA even for three hours. However, treatment with SDS and chloroform destroyed the brucellaphages in 15 min (Fig. 7). While comparing the effect of trypsin and RNAse with Proteinase K and Lysozyme, significant difference (p=0.00) in the phage viability was found. Similarly, significant difference (p=0.00) was observed in phage viability when treated with NS and EDTA in comparison with SDS and chloroform.

Figure 6. Pictorial representation of effect of enzymes on stability of brucellaphages. Phage suspensions were incubated in Proteinase K, Lysozyme, Trypsin, and RNAse for time durations of 15, 30- and 60-min. Post treatment viability was assessed through double agar overlay technique. Phage titers (PFU/mL) (y-axis) were plotted against effect of different enzymes (x-axis) for phage isolate 1 (BaP1), phage isolate 2 (BaP2) and positive control Tbilisi phage (Tb).
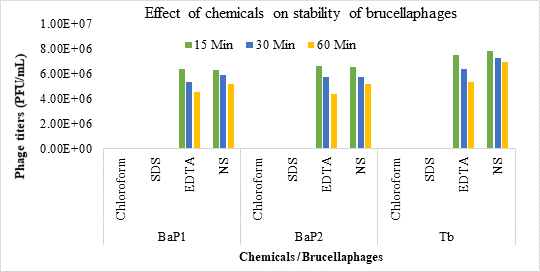
Figure 7. Pictorial representation of effect of chemicals on stability of brucellaphages. Phage suspensions were incubated in SDS, EDTA, NS, and chloroform for time durations of 15, 30- and 60-min. Post treatment viability was assessed through double agar overlay technique. Phage titers (PFU/mL) (y-axis) were plotted against effect of different chemicals (x-axis) for phage isolate 1 (BaP1), phage isolate 2 (BaP2) and positive control Tbilisi phage (Tb).
DISCUSSION
Tb is a reference brucellaphage virus that is used all over the world as standard. It is a typical brucellaphage with respect to its morphology, host range and resistance to physical and chemical agents. It can replicate in almost all B. abortus strains included vaccine as well as field strains (Zhu et al., 2009; Hammerl et al., 2017). In the present study, Tb phage produced pinpoint, round and clear plaques of 0.5 mm diameter but reached maximum of 2.0 mm after 48 hr of incubation with B. abortus strain S99. Plaque morphology of present study brucellaphages BaP1 and BaP2 was same as of Tb phage i.e. pinpoint, round and clear plaques of 0.5 mm diameter. Chachra et al. (2012); Pandey et al. (2013); Gupta and Saxena (2017 a, b) and Saxena and Raj (2018) also found circular and clear plaques of 0.1-3.0 mm diameter of brucellaphages in their studies. Adsorption time determined for present study phages corresponds to Antadze et al. (2017) findings. Broad spectrum lytic activity of isolated brucellaphages i.e. BaP1 and BaP2 indicated that they could not lyse other heterogeneous bacterial species. Similar findings were demonstrated by Pandey et al. (2013) that their brucellaphage could not lyse any of the heterologous bacteria. Prajapati et al. (2014) also found lytic activity of phage against B. abortus strain 99, S19, and 544 as well as B. melitensis Rev 1 and B. suis 1330. But this phage did not lyse any of the heterogeneous bacterial species. However, lytic spots positive for Staphylococcus and Bacillus in our study, might be due to ‘Lysis from without’ as replication of phages inside them has not been observed in plaque assay. Lysis from without is a phenomenon in which death of bacterial cells is caused by Lysozyme like enzymes of high concentration of adsorbed phage particles resulted in collapse of the cell wall. So, interpretation of lysis from without effect is quite difficult and non-specific than plaques formation which explicitly demonstrated phage infection (Hammerl et al., 2017). Hence, these results ascertained that brucellaphage is specific for Brucella and not to other gram positive or negative bacteria, which is advantageous that other microflora in the body will not be disturbed.
In this study, physicochemical characterization of brucellaphages (BaP1 and BaP2) showed that bacteriophages for B. abortus can withstand temperature up to 70oC and had stability between pH 7 to 9. However, acidic pH of 2 to 4 decreased the phage viability in one-hour exposure. Chachra et al. (2012) reported similar findings that at high temperature of 70oC and 3 pH, phage survivability rate was 24.08%. But at pH 7 and 9, brucellaphage had 100% survival ability, when exposed for one hour. Chachra et al. (2012); Pandey et al. (2013) and Gupta and Saxena (2017b) studies are also in agreement to our findings for physicochemical characteristics of brucellaphages. Evaluation of thermal stability and optimization of pH range of brucellaphages are helpful to standardize the phage therapy and its industrial use such as in pasteurized dairy products. Extreme temperature resistance of brucellaphages is beneficial to keep them working in harsh field conditions.
Increasing interest of using bacteriophages to control the diseases entails exploring the favorable long-term storage temperature for bacteriophages. Storage stability of tested BaP1 and BaP2 phages depicted that they had maximum stability at 4oC as they retain their viability up to the 12 months. Although, freezing temperatures were not revealed to be suitable for storage of phages as they lost their viability to 100% after 12-month period at -80oC and 50% at -20oC. There are several records of bacteriophages stored at 4oC maintaining their high titer even after 30 years (Fortier and Moineau, 2009; Merabishvili et al., 2009; Weber-Dąbrowska et al., 2016; Alvi et al., 2018). The decrease in phage titer for long term storage might be due to non-availability of host system (host bacterium and media) or absence of suitable cryopreservatives. Freezing with glycerol was not as effective as preservation at 4oC. It was evident that some bacteriophages form aggregates when stored in high titers at low temperatures which can impact their activity and diluting them promotes a more rapid decrease in their titer (Carlson, 2005; Serwer et al., 2007). So, refrigeration is the most effective and economical means of long-term storage with no fall of phage activity.
BaP1 and BaP2 phages had substantial stability in fluorescent and sunlight and there was no considerable reduction in phage viability after 1-hour exposure. Gradual reduction in brucellaphage titer was observed with increased contact time. With exposure of two hours in sunlight and normal fluorescent light, reduction in phage viability was recorded to 32% and 7%, respectively. However, UV light killed the brucellaphage within 15 min of exposure. Effects of normal, sun and UV lights in our study are similar to the findings of Chachra et al. (2012) who reported that brucellaphage was inactivated within 15 min exposure to UV light, but there was no considerable effect of normal fluorescent light and sunlight within 15 min exposure. However, over a 90 min exposure of phage at room temperature with fluorescent and sunlight, phage survival gradually decreased to 96.17% and 73%, respectively. Pandey at al. (2013) also observed that UV light exposure of phage inactivated it within 3 min and direct sunlight also reduced the phage titer to 93.99% gradually within 3 hr of exposure. Thus, it is perceived that there is a need to keep check on phage survival with contact time, if use in environmental application as sunlight would have some reduction effect on survival of brucellaphage.
BaP1 and BaP2 phages showed stability with some enzymes and reagents but they are vulnerable to other enzymes and chemicals. Our results correlated with Gupta and Saxena (2017), who stated that when brucellaphage was incubated with lysozyme it was completely inactivated in one hour but no detectable change was found when treated with RNase enzyme for 120 min. In the same way, when brucellaphage was treated with NSS and EDTA, even for three hours, there was no effect on the viability of phage, but SDS and chloroform treatment destroyed the brucellaphage in 15 min. Pandey et al. (2013) also observed phage stability with RNase treatment up to 3 hr and inactivation of phage after treatment with chloroform, SDS and lysozyme.
Conclusion: In conclusion, this study will corroborate to standardize the various cost-effective applications of brucellaphages such as phage therapy and specific decontamination of B. abortus in livestock farms. Molecular and genetic characterization as well as electron microscopy of isolated brucellaphages is suggested in future studies before in-vitro and in-vivo experimental evaluation. Moreover, having developed the strategic procedures in the laboratory in Pakistan, and having successfully isolated indigenous isolates of brucellaphages (BaP1 and BaP2), we are equipped to isolate bacteriophages against field strains of B. abortus for effective bio-control of bovine brucellosis in the future.
Authors’ contribution: AAS conceived and designed the study. MR and WS reviewed the study project for final approval. AYS executed the experiments and analyzed the study results. ZA supported for the technical assistance. MM helped in execution of research experiments. AYS and AAS critically revised the manuscript for important intellectual contents and final version was approved by all authors.
Acknowledgments: The authors gratefully acknowledge the financial support provided by Higher Education Commission (HEC) project # 6782/Punjab/NRPU/R&D/HEC/2016 under National Research Program for Universities, Pakistan and Denis Tremblay, (University Laval, Quebec City, QC, Canada) for the provision of positive control phage.
REFERENCES
- Abubakar, M., M. Mansoor and M.J. Arshed (2012). Bovine brucellosis: old and new concepts with Pakistan perspective. Pak. Vet. J. 32(2): 147-155.
- Alvi, I.A., M. Asif, R. Tabassum, Z. Abbas and S. ur Rehman (2018). Storage of bacteriophages at 4oC leads to no loss in their titer after one year. Pak. J. Zool. 50(6): 2395-2398.
- Antadze, I., Dadunashvili, T. Burbutashvili, S. Gunia, N. Balarjishvili, E. Tevdoradze, T. Pataridze, R.J. Obiso, S. Hagius, P. Elzer and M. Kutateladze (2017). Diversity of phage-host specificity in Brucella Phage. J. Bacteriol. Mycol. 4(2): 1-8.
- Carlson, K. (2005). Working with bacteriophages: Common techniques and methodological approaches. Vol. 1. CRC Press; Boca Raton. pp. 439-490.
- Chachra, D., H. Kaur, M. Chandra and H. Saxena (2012). Isolation, electron microscopy and physicochemical characterization of a brucellaphage against Brucella abortus vaccine strain S19. Int. J. Microbiol.10: 1-7.
- Farlow, J., A.A. Filippov, K.V. Sergueev, J. Hang, A. Kotorashvili and M.P. Nikolich (2014). Comparative whole genome analysis of six diagnostic brucellaphages. Gene. 541: 115-122. doi: 10.1016/j.gene.2014.01.018
- Filippov, A.A., K.V. Sergueev and M.P. Nikolich (2013). Bacteriophages against biothreat bacteria: diagnostic, environmental and therapeutic applications. J. Bioterr. Biodef. S3: 1-8.
- Flores, V., A. López-Merino, G. Mendoza-Hernandez and G. Guarneros (2012). Comparative genomic analysis of two brucellaphages of distant origins. Genomics. 99: 233-240.
- Fortier, L.C. and S. Moineau (2009). Phage production and maintenance of stocks, including expected stock lifetimes. Methods. Mol. Biol. 501: 203-219.
- Gopalaiah, H. (2013). Bacteriophage as antimicrobial agents: A milestone. J. Indian Acad. Oral Med. Radiol. 25 (1): 40-41.
- Gupta, V. and H.M. Saxena (2017a). A new bacteriophage based luminescence assay for diagnosis of brucellosis. Indian J. Exp. Biol. 55: 296-302.
- Gupta, V. and H.M. Saxena (2017b). Isolation and characterization of BpL1, a broad acting lytic bacteriophage against Brucella. Int. J. Curr. Microbiol. App. Sci. 6(11): 2486-2496.
- Hammerl, J.A., S. Al Dahouk, K. Nockler, C. Gollner, B. Appel and S. Hertwig (2014). F1 and tbilisi are closely related brucellaphages exhibiting some distinct nucleotide variations which determine the host specificity. Genome Announc. 2(1): e01250-13.
- Hammerl, J.A., C. Gollner, C. Jackel, H.C. Scholz, K. Nockler, J. Reetz, S. Al Dahouk and S. Hertwig (2017). Genetic diversity of Brucella reference and non-reference phages and its impact on Brucella-typing. Front. Microbiol. 8(408): 1-12.
- Hamza, A., S. Perveen, Z. Abbas and S.U. Rehman (2016). The lytic SA phage demonstrates bactericidal activity against mastitis causing Staphylococcus aureus. Open Life Sci. 11(1): 39-45.
- Hyman, P. and S.T. Abedon (2010). Bacteriophage host range and bacterial resistance. Adv. Appl. Microbiol. 70: 217-248.
- Jain, L., M. Rawat, A. Prajapati, A.K. Tiwari, B. Kumar, V. Chaturvedi, H. Saxena, S. Ramakrishnan, J. Kumar and P. Kerketta (2015). Protective immune-response of aluminium hydroxide gel adjuvanted phage lysate of Brucella abortus S19 in mice against direct virulent challenge with abortus 544. Biologicals. 43: 369-376.
- Manish, K., C. Puran, C. Rajesh, R. Teena and K. Sunil (2013). Brucellosis: an updated review of the disease. Indian J. Anim. Sci. 83(1): 3-16.
- Merabishvili, M., J.P. Pirnay, G. Verbeken, and N. Chanishvili, M. Tediashvili, N. Lashkhi and L. van Parys (2009). Quality-controlled small-scale production of a well-defined bacteriophage cocktail for use in human clinical trials. PLoS One. 4(3): e4944.
- Moghadam, M.T., N. Amirmozafari, A. Shariati, M. Hallajzadeh, S. Mirkalantari, A. Khoshbayan and F.M. Jazi (2020). How phages overcome the challenges of drug resistant bacteria in clinical infections. Infect. Drug. Resist. 13: 45-61.
- OIE (2018). Chapter 1.1.4 Biosafety and Biosecurity: Standard for managing biological risk in the veterinary diagnostic laboratories and animal facilities. Manual of Diagnostic Tests and Vaccines for Terrestrial Animals. https://www.oie.int/fileadmin/Home/eng/Health_standards/tahm/1.01.04_BIOSAFETY_BIOSECURITY.pdf
- OIE (2019). Chapter 3.1.4. Brucellosis (Brucella abortus, melitensisand B. suis) (infection with B. abortus, B. melitensis and B. suis). Manual of Diagnostic Tests and Vaccines for Terrestrial Animals. http://www.oie.int/fileadmin/Home/eng/Health_standards/tahm/3.01.04_BRUCELLOSIS.pdf
- Pandey, S., D. Chachra, M. Chandra and H. Saxena (2013). Protein profiling and physico-chemical characterization of an isolated phage against Brucella abortus strain 19. Afr. J. Microbiol. Res. 7: 1233-1238.
- Prajapati, A., D. Ramchandran, H. Verma, M. Abbas and M. Rawat (2014). Therapeutic efficacy of Brucella phage against Brucella abortus in mice model. Vet. World. 7: 34-37.
- Prestel, E., S. Salamitou and M.S. DuBow (2008). An examination of the bacteriophages and bacteria of the Namib desert. J. Microbiol. 46: 364-372.
- Principi, N., E. Silvestri and S. Esposito (2019). Advantages and limitations of bacteriophages for the treatment of bacterial infections. Front. Pharmacol. 10:(513). Doi: 10.3389/fphar.2019.00513
- Rahman, M.S., M.J. Uddin, J.H. Park, J.S. Chae, M.B. Rahman and M.A. Islam (2006). A short history of brucellosis: special emphasis in Bangladesh. Bangladesh J. Vet. Med. 4: 1-6.
- Saxena, H.M. and S. Raj (2018). A novel immunotherapy of Brucellosis in cows monitored non invasively through a specific biomarker. PLoS. Negl. Trop. Dis. 12(4): 1-12.
- Sergueev, K.V., A.A. Filippov and M.P. Nikolich (2017). Highly sensitive bacteriophage-based detection of Brucella abortus in mixed culture and spiked blood. Viruses. 9(144): 1-17.
- Serwer, P., S.J. Hayes and K. Lieman (2007). Aggregates of bacteriophage 0305φ8-36 seed future growth. Virol. J. 4(1):131.
- Shang, D., X. Donglou and Y. Jiming (2002). Epidemiology and control of brucellosis in China. Vet. Microbiol. 90: 165-182.
- Srinivasiah, S., J. Bhavsar, K. Thapar, M. Liles, T. Schoenfeld and K.E. Wommack (2008). Phages across the biosphere: contrasts of viruses in soil and aquatic environments. Res. J. Microbiol. 159: 349-357.
- Tevdoradze, E., J. Farlow, A. Kotorashvili, N. Skhirtladze, I. Antadze, S. Gunia, N. Balarjishvili, L. Kvachadze and M. Kutateladze (2015). Whole genome sequence comparison of ten diagnostic brucellaphages propagated on two Brucella abortus Virol. J. 12(66): 1-11.
- Texas (2011). Protocol: Phage enrichments. Center for Phage Technology Texas A&M University, College Station, TX 77843. https://cpt.tamu.edu/wordpress/wp-content/uploads/2011/12/Phage-enrichments-07-12-2011.pdf (Accessed on: 21 April 2016).
- Weber-Dąbrowska, B., E. Jonczyk-Matysiak, M. Zaczek, M. Lobocka, M. Lusiak-Szelachowska and A. Gorski (2016). Bacteriophage procurement for therapeutic purposes. Front. Microbiol. 7: 1-14.
- Wommack, K.E. and P.R. Colwell (2000). Virioplankton: viruses in aquatic ecosystems. Microbiol. Mol. Biol. Rev. 64: 69-114.
- Yang, H., L. Liang, S. Lin and S. Jia (2010). Isolation and characterization of a virulent bacteriophage AB1 of Acinetobacter baumannii, BMC. Microbiol. 10. http://bmcmicrobiol.biomedcentral.com/ articles/10.1186/1471-2180-10-131
- Zhu, C.Z., H.Y. Xiong, J. Han, B.Y. Cui, D.R. Piao, Y.F. Li, H. Jiang, Q. Ren, X.Y. Ma and Y.M. Chai (2009). Molecular characterization of Tb, a new approach for an ancient brucellaphage. Int. J. Mol. Sci. 10(7): 2999-3011.
|





